de | fr | en Druckansicht ![]()
3R-INFO-BULLETIN 27
September 2004
The Authors

Dr. Patrick Guerin (left) is Director of Research in Animal Physiology, Institute of Zoology, University of Neuchâtel. He is the head of a research group on sensory physiology and ecophysiology of bloodsucking arthropod vectors of diseases. His research covers chemical stimuli from hosts that play a key role in host selection by these ectoparasites.
Dr. Thomas Kröber (right) is a PostDoc in this research group and focuses his investigations on factors guiding host attachment and feeding in ticks. His primary goal is the development of experimental methods to investigate these vital processes in ticks.
Current addresses:
Dr. Patrick Guerin
patrick.guerin@unine.ch
Dr. Thomas Kröber
thomas.kroeber@unine.ch
Dep. of Animal Physiologie
Institute of Zoology
University of Neuchâtel
Rue Emile Argand 11
CH-2007 Neuchâtel
Switzerland
Editor
Peter Maier, Scientific Adviser of the 3R Research Foundation
The tick blood meal: From a living animal or from a silicone membrane?
Animals as hosts for ticks
Animal husbandry could not be practised over large areas of the planet without acaricides. This persistent reliance on pesticides has led to the development of resistance in ticks against the major classes of acaricide treatments. There is a continual requirement for new types of molecules to target physiological processes that are crucial to tick survival. The development of animal health products against ticks[*] requires hundreds of cattle, dogs, rabbits and gerbils for in vivo trials with acaricides, placing the annual worldwide use of animals in acaricide research in the tens of thousands. Small mammals used in such trials may suffer from skin inflammation and anaemia, and may be submitted to restrictions by the Elizabethan collar inhibiting cleaning behaviour. For controlled studies, dogs have to be kept in small cages, cattle are kept in isolation, in climatic boxes, where their movement is confined. Apart from the ethical aspects of using experimental animals, the costs of maintaining suitable hosts for ticks are high.
In vitro feeding: outwitting the ticks
Tick control on animals is achieved either through contact with a product applied either topically or orally. Ideally, an in vitro assay should permit both the assessment of products that either affect a tick's capacity to attach for a blood meal, or that restrict feeding once the tick has started to take blood. When ticks start to attach to the skin, they penetrate the uppermost keratin rich stratum corneum with outward lacerating movements of their cutting mouthparts. Strong retrograde food canal denticles anchor the tick in the skin (Fig. 1) allowing the cutting mouthparts to move deeper until the corium containing blood vessels is reached. The artificial feeding unit, that we have developed, has a silicone membrane that replaces host skin and provides the tick with a similar perch over blood (Fig. 2).

Fig. 1: Ixodes ricinus female attached to the silicone membrane. Half of the 500 μm long food canal has pierced the membrane. (Photo: M. Vlimant)
The feeding unit consists of a silicone membrane reinforced with cellulose rayon glued across one end of a piece of acrylic glass tubing (44 mm high and 26 mm i.d.). The membrane is a modified version of one developed in a previous 3R project (No. 10-88)1,2. We rendered the membrane softer to facilitate feeding by the European tick, Ixodes ricinus. The new membrane allows ticks with shorter mouthparts (500 μm in the case of female I. ricinus; Fig. 1) to be accommodated and the mouthparts of ticks to be withdrawn and reattach elsewhere. The previous penetration site closes by elastic retraction forces in the membrane preventing leaking of blood into the unit. The feeding units are set up in six well plates (Fig. 2) where each is equipped with an outer ring that limits the depth to which it sinks into the 3.5 ml of blood applied per well. Tick feeding proceeds at body temperature on a support in a water bath. Bovine blood, collected weekly from an abattoir, is manually defibrinated and supplemented with glucose, an antibiotic and ATP (as a feeding stimulus). Blood is exchanged twice daily.

Fig. 2a: Six-well plate with feeding units over blood.
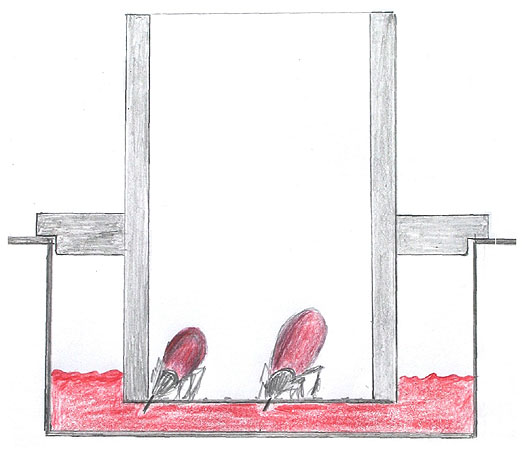
Fig. 2b: Schema of a section through one feeding unit depicting the ticks feeding on blood through the membrane.
Attachment by ticks at predilection sites on hosts is preceded by a behavioural sequence that depends on the presence of an appropriate array of mechanical, olfactory and contact chemostimuli3. We achieved a 75-95% attachment rate by I. ricinus in these feeding units by applying a combination of chemical and mechanical stimuli. A cow hair extract was applied to the membrane in addition to mosquito netting and a layer of cow hair (Figs. 2 and 3).
Testing an acaricide in vitro
Fipronil, an odourless phenyl pyrazole, is toxic to a broad range of arthropods via contact or ingestion. This acaricide disrupts ion flow by interacting with GABA-gated chloride channels of the CNS causing hyperexcitation. The effect of different doses of fipronil dissolved in the bovine blood on I. ricinus mortality was assessed over 9 days in our feeding units. Fipronil killed all females within two days at 10 μg/ml of blood, at 1 μg/ml no females survived longer than four days, and at 0.1 μg/ml all females were killed by day 7 (Fig 4). These are similar to the doses of fipronil required to protect companion animals against ticks. At the lowest doses of 0.01 and 0.001 μg fipronil/ml female ticks survive but their feeding activity is reduced by 35% and reproduction is inhibited. Females feeding on 0.001 μg fipronil/ml laid eggs but none of these hatched.

Fig. 3: Feeding unit with partially engorged female Ixodes ricinus sucking bovine blood for over a week. Less engorged females (see blue arrow) can readily reattach to complete the blood meal.
Since fipronil, like other acaricides, is also applied to animals as a spot-on in the fur, we tested the contact effects of this acaricide in our feeding units by applying it to the surface of the silicone membrane before the ticks were introduced to the feeding units. Fipronil applied in this manner inhibited feeding and strongly affected tick survival: mortality reached 69 % at 10 ng fipronil/cm2 and 100 % at 1 μg fipronil/cm2 within 30 h (controls 19 %, P 0.001 and 0.0001, respectively, Fisher's exact test).
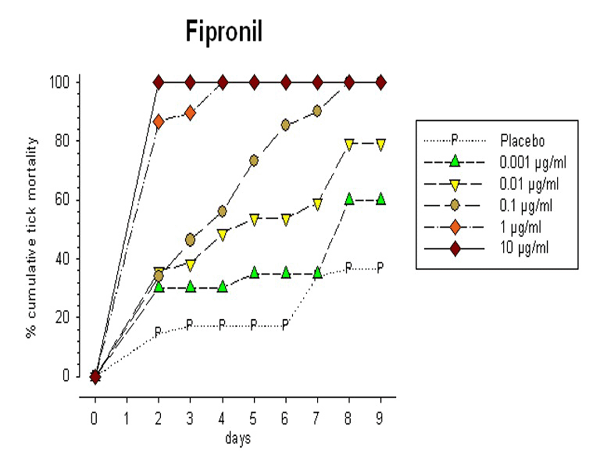
Fig. 4: Cumulative mortality of Ixodes ricinus females feeding on bovine blood through a silicone membrane with dimethylsulfoxide (DMSO) added (placebo) and with increasing doses of fipronil in DMSO.
Advantages of the in vitro feeding assay
In vivo trials with animals as hosts require repetitions with 10 to 20 animals due to the inherent variation between them. Furthermore, large amounts of products and ticks are needed for such test. By contrast, our in vitro assay requires approximately one hundred fold less of the test product and only about 40 ticks are required per dose. Survival curves calculated over the different doses of fipronil in different feeding experiments showed that the observed effects were significant, were obtained within 5-6 days and highly reproducible. In addition, this in vitro assay permits setting up more standardised conditions since the placebo, a reference acaricide and test products can be tested in blood from the same donor animal. Together these reasons suggest that the in vitro feeding assay for hard ticks is preferable to in vivo screening trials on animals.
Published updated version of this Bulletin 27/2007 (PDF)
References:
- Kuhnert F, Diehl PA and Guerin PM, The life-cycle of the bont tick Amblyomma hebraeum in vitro. Int J Parasitol 25, 887-896 (1995).
- Kuhnert F, Feeding of hard ticks in vitro: New perspectives for rearing and for the identification of systemic acaricides. ALTEX 13, 76-87 (1996).
- Guerin PM, Kröber T, McMahon C, Guerenstein P, Grenacher S, Vlimant M, Diehl PA, Steullet P and Syed Z, Chemosensory and behavioural adaptations of ectoparasitic arthropods. Novo Acta Leopoldina 83, 213-229 (2000).
| [*] | The tick blood mealThe prevention of tick bites in animals reduces tick-transmitted diseases such as anaplasmosis, babesiosis, theileriosis and heartwater disease. The duration of the tick's blood meal takes 2 to 14 days depending on whether it is a larva, nymph or adult. During the first days of feeding, only small amounts of blood are imbibed. However, this a period during which the tick undergoes a variety of physiological changes to accommodate the blood meal in females. Numerous physiologically active agents are injected by the parasite into the feeding lesion inducing strong inflammatory, vasodilatory and immunological responses by the host. The tick's engorgement occurs during the last 24 hours of feeding during which the females imbibe 2 to 8 times as much blood as they finally gain in weight and multiply their body mass by as much as 100 fold with protein and lipid rich nutrient for the production of thousands of eggs. Males do not engorge themselves with blood. In some ticks (e.g. the European tick, Ixodes ricinus), every life stage feeds on a different host: larvae and nymphs mainly on small vertebrates and females on sheep, deer and other larger mammals. As man is occasionally parasitized, this contributes to this tick's role as a vector of spring-summer meningoencephalitis and bacterial (Lyme borelliosis) diseases. |
| Letzte Änderung: 30.01.2008 |